Introduction
Phagocytosis is one of the oxygen depending processes in organisms. During the unspecific immunological defense, the activity of pentose phosphate pathway is dramatically increased forming NADPH. NADPH is needed to reduce the oxygen which is bonded to membrane-based cytochromes.
Therefore, the oxygen demand is strongly increased (respiratory burst). During this process, oxygen is converted into superoxide anions, hydrogen peroxide, monomolecular oxygen and hydroxyl c radicals by means of several kinds of phagocytic cells (e.g. neutrophil, eosinophil and basophil leucocytes, macrophages). These extracellular highly reactive oxygen species (ROS) cause many biological effects such as destruction of bacterial cells, parasites and tumor cells, promoting inflammation and modulating the immune reaction (Fig. 1).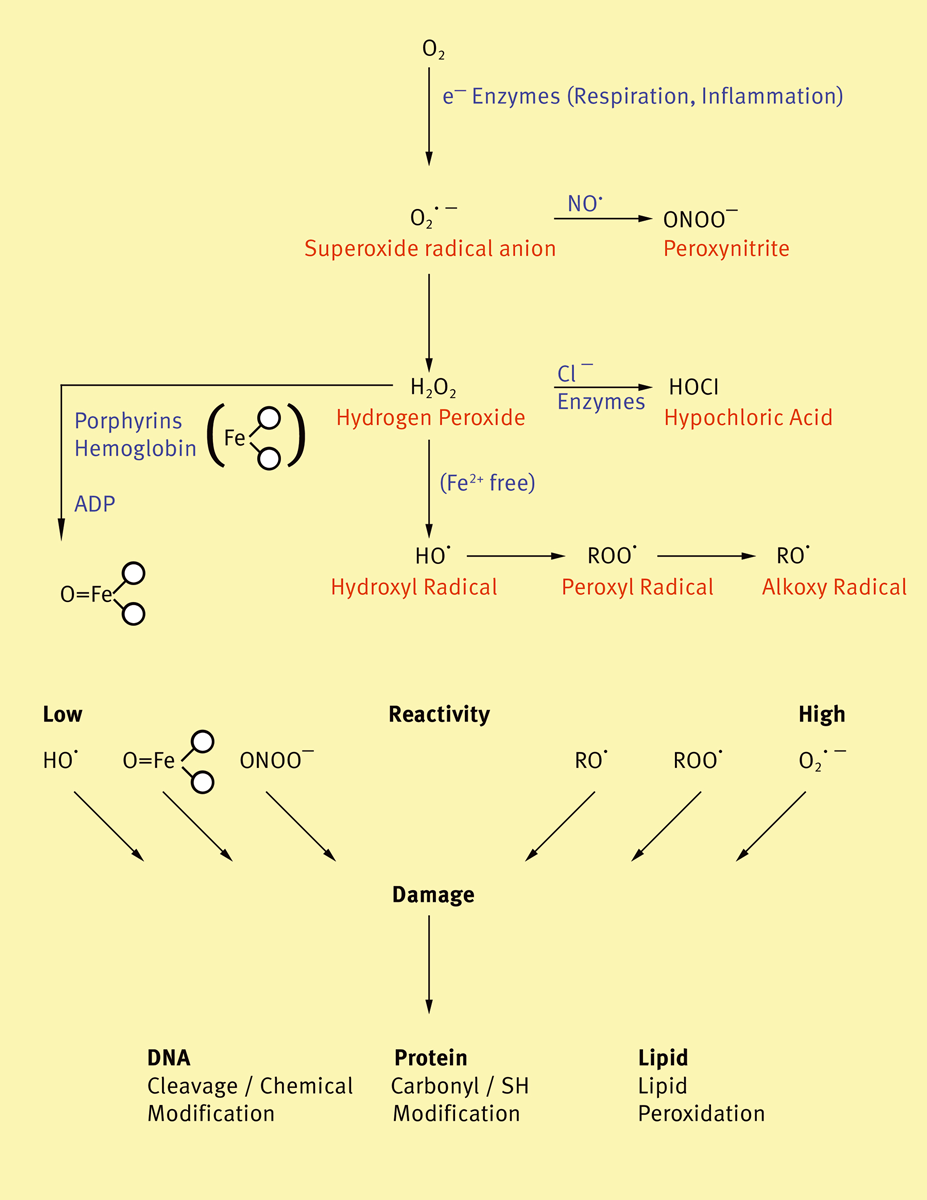
The generation processes of reactive oxygen species can be monitored using luminescence analysis. Measurements of chemiluminescence (CL) are highly sensitive and specific, owing the possibility to investigate the different kinds of reactive oxygen species simultaneously (HO, O2, H2O2, 1O2). Because of the very weak native luminescence phenomena, luminol or lucigenin dependent chemiluminescence has been used frequently for the detection of superoxide radical anions in biological systems. Luminol reacts in its univalently oxidized form and lucigenin reacts in its univalently reduced form with O2. In both cases, light production depends on the formation of an unstable endoperoxide or dioxetane, which decomposes to an electronically excited product. This product releases a photon as it falls to the ground state.
In this application note we show the measurement of ROS generation in the yeast Candida albicans (C. albicans) using lucigenin. In C. albicans, mitochondria are capable of generating extracellular released ROS. The effect of terbinafine, a considered free radical interceptor, on ROS formation is investigated.
Materials & Methods
The following strains of Candida were used for the measurement of cellular luminescence: C.albicans, C.tropicalis, C.guilliermondii, C.glabrata, C.parapsilosis.
The strains of fungi were cultured 24 h on Sabouraud-Glucose-Agar in the presence of penicillin and gentamicin. Yeast suspensions were prepared in RPMI-1640 medium with concentrations of 107 to 1011 blastospores/mL or in isotonic NaCl solution with the same concentration. 200 µL of several yeast suspensions were used. Candida cells at a concentration of 1x108 cells/mL were incubated in saline with terbinafine at concentrations of 1 µg/mL, 10 µg/mL and 100 µg/mL dissolved in DMSO. Incubation was performed for 10 min and 60 min at 25°C in a shaking incubator. The duration was restricted to exclude effects of terbinafine on cell number and growth. Lucigenin solution was prepared in PBS at a concentration of 10-4 mol/L and added into each well at 30°C or 37°C by an injection pump.
Instrument settings
|
Omega series |
CLARIOstar |
PHERAstar FS |
|
|
Detection mode |
Luminescence |
||
|
Method |
Plate mode kinetic |
||
|
Optic settings |
lens |
lens |
Luminescence |
|
Measurement time per data |
1 second |
||
|
Measurement time per cycle |
60 seconds |
||
|
Number of cycles |
20 |
||
Samples were taken from yeast strains at different cell numbers and for a fixed cell number with varying terbinafine concentration. The calculation was performed using the average counts per minute (cpm) over measurement periods of 20 min.
Results & Discussion
The results of lucigenin dependent chemiluminescence measured in suspensions of Candida albicans, Candida glabrata, Candida guilliermondii, Candida parapsilosis, Candida tropicalis are shown in Table 1.
Table 1: Comparison of different yeast species.
|
Candida albicans |
Candida guilliermondi |
Candida parapsilosis |
Candida tropicalis |
Candida glabrata |
Trichosporon capitatum |
|
|
cells |
RLU |
RLU |
RLU |
RLU |
RLU |
RLU |
|
E+7/mL |
0 |
0 |
2 |
0 |
0 |
19 |
|
E+8/mL |
0 |
0 |
4 |
0 |
7 |
94 |
|
E+9/mL |
2 |
12 |
19 |
25 |
35 |
101 |
|
E+10/mL |
30 |
48 |
61 |
90 |
126 |
124 |
|
E+11/mL |
64 |
122 |
156 |
102 |
189 |
324 |
|
|
r = 0.91 |
r = 0.96 |
r = 0.960 |
r = 0.74 |
r = 0.84 |
r = 0.970 |
(RPMI-medium, T=30°C, Lucigenin-solution 100 µL)
It was notable that the generation of reactive oxygen species in C. albicans could not be detected at concentrations of 107 to 109 blastospores/mL. Detect-able CL values were measured at ≥ 1010 blastospores/mL. The reproducibility of the results could be demonstrated by inter assay variation as well as linear correlation (r=0.91 resp. 0.98) of the concentration of blastospores and the relative light units (RLU). The linear and direct proportion of the ROS generation and blastospore concentration could be confirmed due to the calculated correlation coefficients. The reproducibility of the results has been demonstrated by inter assay variation on several days (data not shown). After 10 min of incubation with terbinafine, a dose-dependent inhibition of ROS generation was already seen (Fig. 2).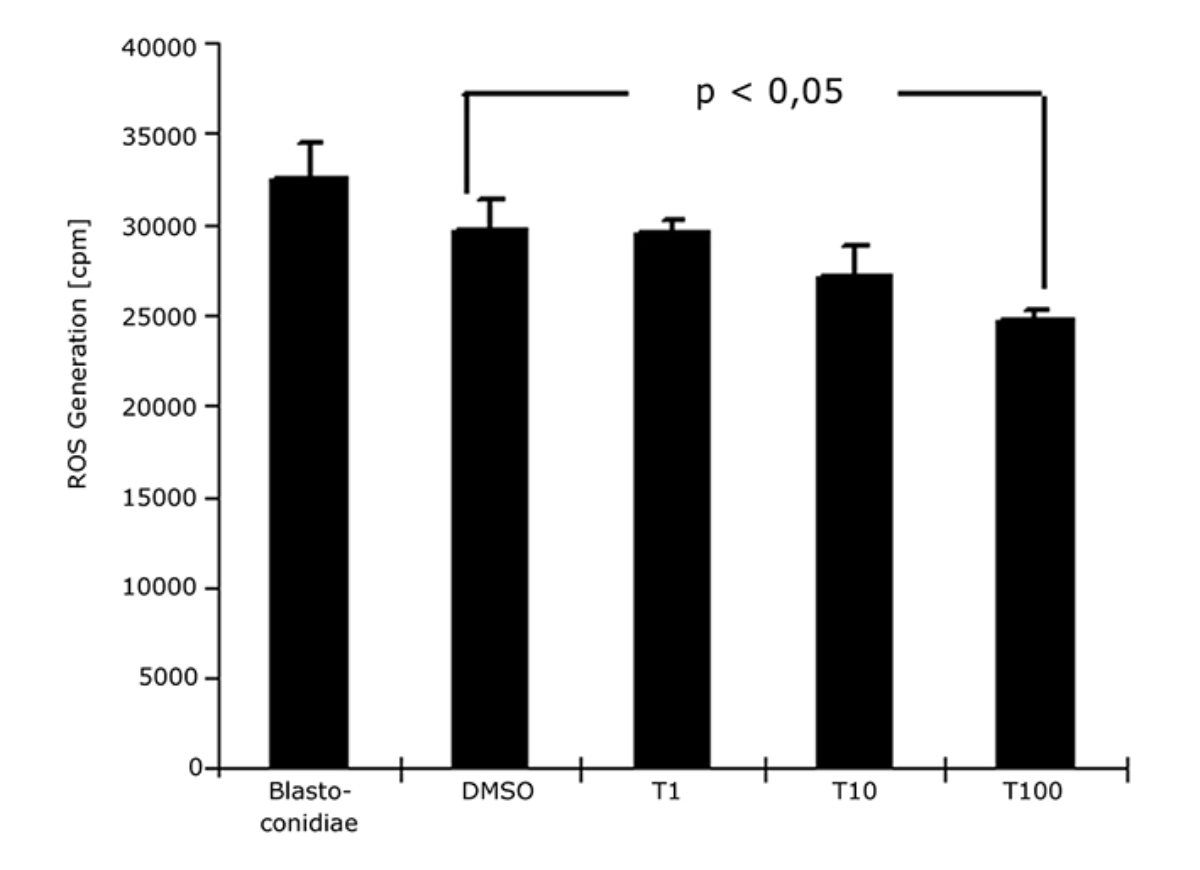
DMSO itself, as the soluble reagent, revealed anti-oxidant properties. However, the inhibition of ROS generation was considerably dependent on the used terbinafine concentration. At a terbinafine concentration of 1 µg/mL (T1) the inhibition corresponded to 9.4 % and at 100 µg/mL (T100) to 24 %.
The inhibition was significant for the comparison of the control incubated with DMSO and the highest chosen terbinafine concentration of 100 µg/ml (T100) (p<0.05). After 60 min of incubation with terbinafine the inhibition was amplified (data not shown). The final inhibitory effect was 18 % for 1 µg/mL terbinafine (T1) and 41.1 % for 100 µg/mL terbinafine (T100). The effect was significant for terbinafine concentrations of 10 µg/mL (T10) and 100 µg/mL (T100) (p < 0.05). The dose of 10 µg/mL (1 %) corresponds to the dose applied for topical preparations.
Conclusion
The ability of various candida yeasts and blastomyces to generate ROS can be monitored by means of lucigenin-dependent chemiluminescence. The CL method is sensitive enough to detect very low ROS concentrations produced by several fungi without any stimulation.
This was used to look for terbinafine effects on the ROS formation in C.albicans. The results confirm a terbinafine time- and dose-dependent inhibition of ROS generation by C. albicans. Due to the time-dependency, it is likely that terbinafine not only has radical scavenging properties, but also interacts with the pathway that generates reactive oxygen species. In summary, terbinafine reduces the ability of C. albicans to generate reactive oxygen species.