Introduction
Targeted protein degradation is a novel strategy with the aim to selectively eliminate specific proteins from cells for therapeutic reasons. While this research mostly focused on PROTAC molecules at the beginning, nowadays new target degrader approaches are emerging. One of these is the degradation TAG (dTAG) technology.
dTAG is an innovative approach to target validation. Like PROTACs, this approach uses a heterobifunctional small molecule to establish ternary complex formation between a target protein of interest and an E3 ubiquitin ligase. However, what sets dTAGs apart, is that the binding site for one part of the bifunctional degrader is artificially introduced into the target protein by CRISPR/Cas9. This marks dTAGs as ideal tools for performing target validation studies for dose-dependent effects.
Assay principle
The dTAG protein degradation assay highlighted in this application note utilises a dual luciferase reporter system to measure target protein degradation. For this purpose, one luciferase, firefly luciferase in this case, is used as a house-keeping marker that should not be affected by dTag degradation. The second luciferase, nanoluciferase, is present in its fragmented inactive form HiBiT and fused to the target protein of interest. The dTAG degraders are able to establish a bond between the target protein and a E3 ubiquitin ligase via the introduced dTAG binding motif which forms a ternary complex (Fig. 1). This leads to the ubiquitination and subsequent degradation of the target protein and the fused HiBiT fragment. In order to measure nanoluciferase luminescence, LgBiT is added after degradation. This small nanoluciferase fragment can complete undegraded HiBiT fragments, leading to an active nanoluciferase enzyme and consequent light emission. The ratio of the nanoluciferase/firefly emission is an indicator of the efficacy of the dTAG degrader with lower ratios as a proxy for higher target degradation. 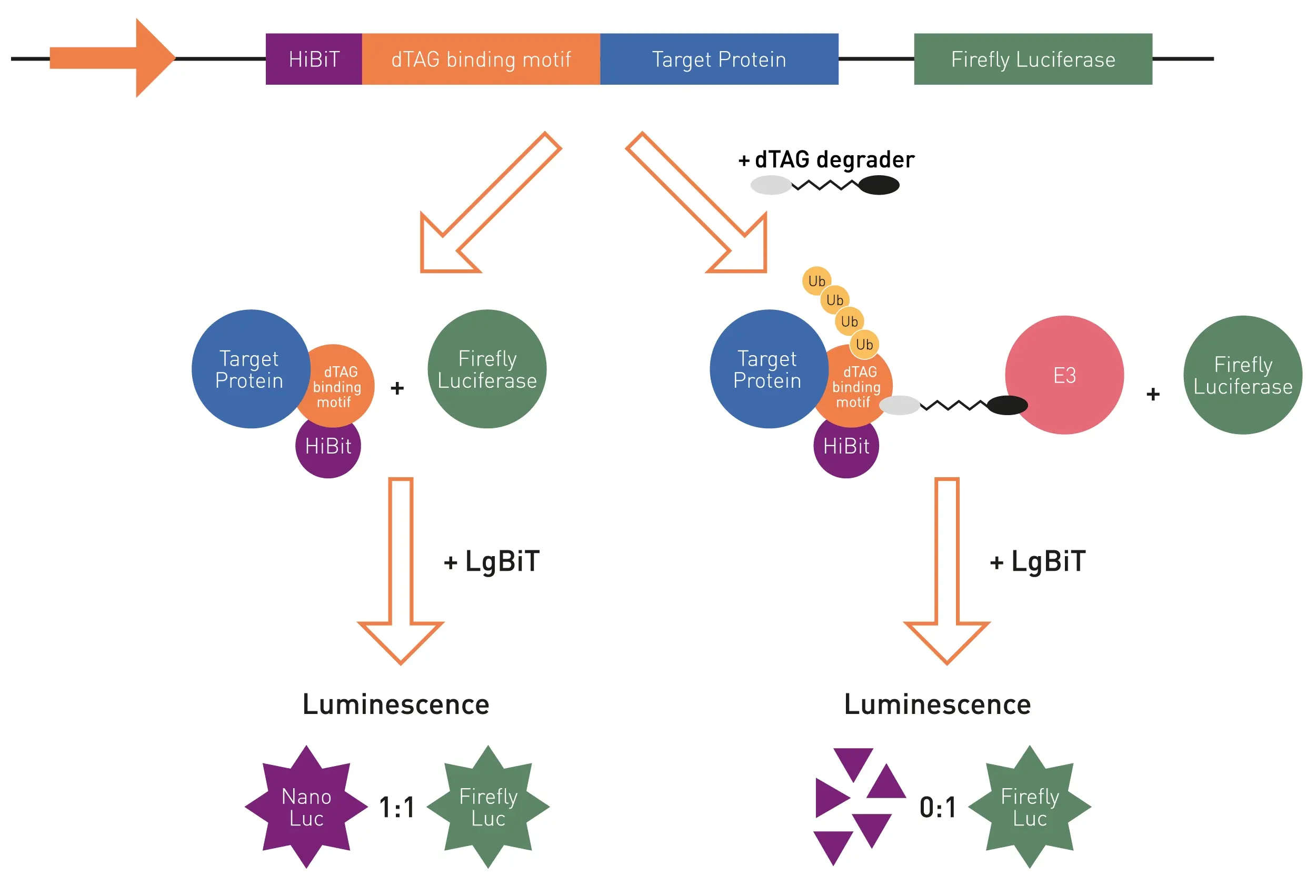
Materials & methods
- 1536-well iSTAR microplate (GT249.008, Bio-Gene)
- PHERAstar FSX (BMG LABTECH)
- pSBTK-HiBiT-dTAG-TargetX-P2A-Fluc-T2A-Puro plasmid (Promega)
- HEK293T cells
- Nano-Glo dual luciferase reporter assay system (N1650, Promega)
Experimental Procedure
dTAG Protein degradation assay
dTAG degraders or DMSO controls were dispensed to the wells of a 1536-well iSTAR microplate. HEK293 cell monolayers stably transfected with the pSBTK-HiBiT-dTAG-TargetX-P2A-Fluc-T2A-Puro plasmid were detached from their culture flask to create a cell suspension. After centrifugation at 1,400 rpm for 4 min, the supernatant was removed, and the cell pellet was re-suspended in OPTIMEM medium containing 2% FBS. 2.5 µL of this cell suspension were added per well to the 1536-well iSTAR plate. The plate was centrifuged briefly and kept at 5% CO2 and 37°C for 4 hours. 2.5 µL OneGlo reagent was added per well and firefly luminescence was read after 10 min incubation and shaking at 300 rpm at the PHERAstar FSX. Afterwards, 2.5 µL StopGlo reagent, 0.025 µL nanoluciferase substrate and 1/100 LgBiT were added per well, the plate was centrifuged for 1 min and nanoluciferase luminescence was read at PHERAstar FSX.
Instrument settings
|
Luminescence, endpoint
|
||
|
Optic settings
|
Filters
|
-
|
|
General settings
|
Integration time
|
0.1 s
|
|
Focal Height
|
13.5 |
|
|
Gain
|
3600 |
|
Results & Discussion
Several dTAG degraders were tested for their ability to induced targeted degradation of HiBiT. For this purpose, increasing concentrations (0.1 to 10,000 nM) of the degraders dTAG-v2, dTAG-47, and dTAG-13 were added to HiBiT-expressing cells in a 1536-well microplate (Figure 2).
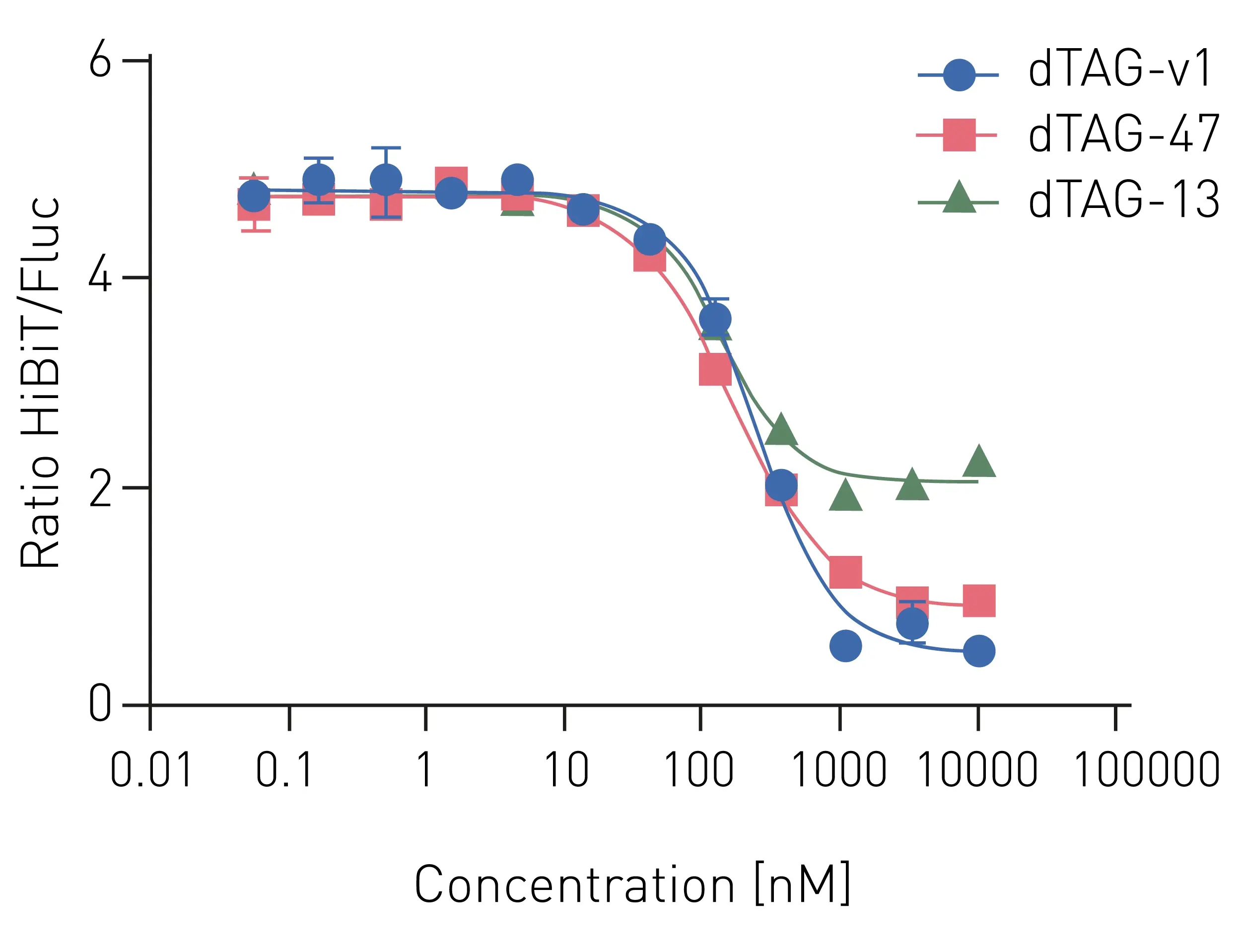
All tested dTAG degraders induced a decrease in nanoluciferase luminescence (i.e., HiBiT degradation) in a concentration-dependent manner. However, different extents of HiBiT degradation could be observed between the dTAG molecules. While dTAG-13 (green curve) lead to an about 50% degradation and decrease of nanoluciferase luminescence, dTAG-v1 (blue curve) proved to be the most potent degrader with a nearly complete degradation of HiBiT.
Data stability of dTAG-v1 degrader data was further characterized in a second experiment (Fig. 3). Here, dTAG-v1 concentrations ranging from 1 to 10 µM were evaluated with 32 replicates in comparison to DMSO controls.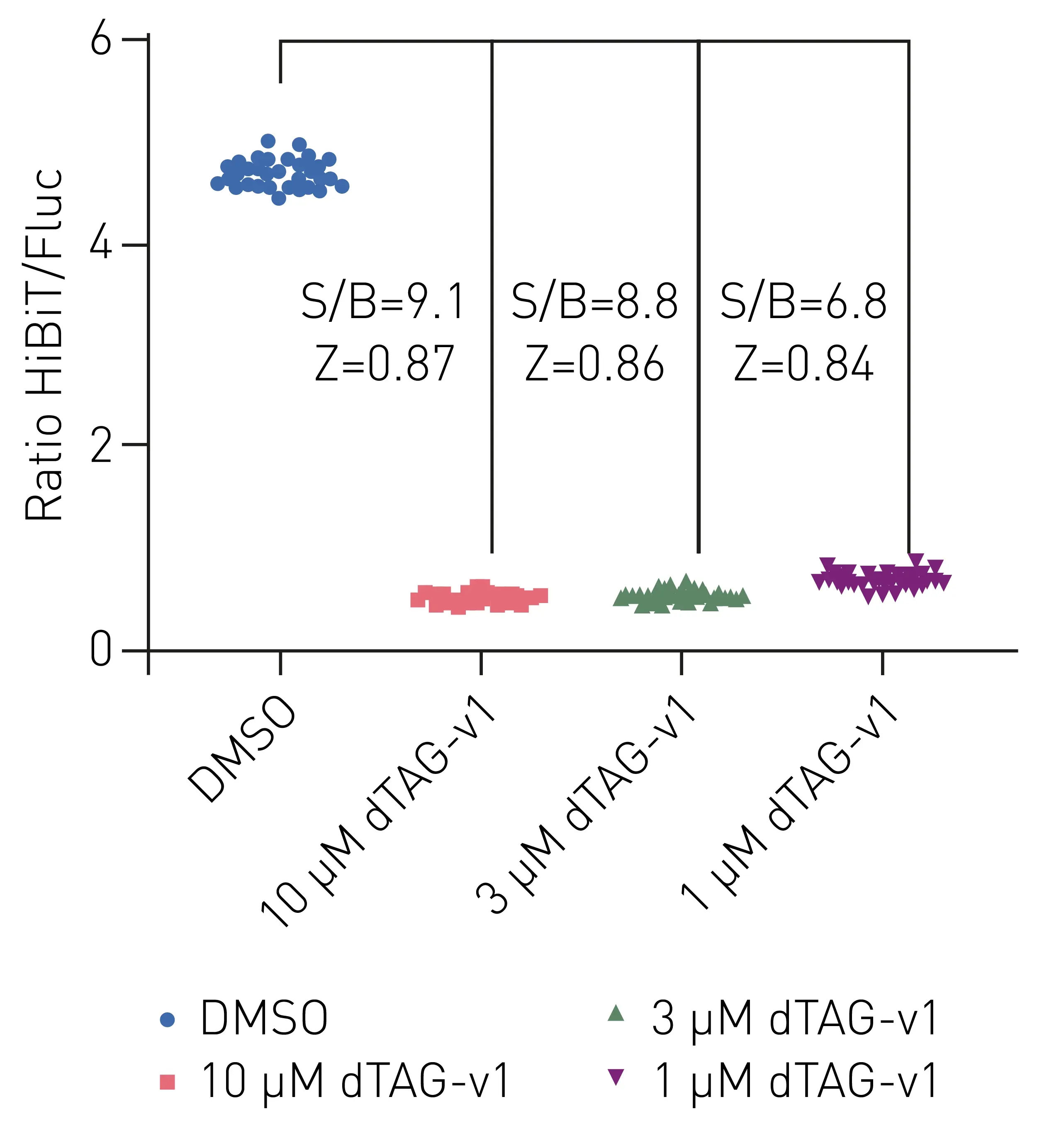
As can be seen in figure 3, all three applied concentrations of dTAG-v1 led to degradation of HiBiT-fusion protein in comparison to DMSO controls. A concentration-dependent decrease of nanoluciferase luminescence signal could be observed as highlighted by the signal to blank ratios, while Z factors of 0.84 to 0.87 depicted good data stability.
Conclusion
Using the dTAG protein degradation assay, the tested dTAG molecules could be successfully employed to induce the targeted degradation of the HiBiT-fusion protein complex. Due to its high sensitivity, fast read times, and downscaling compatibility, the PHERAstar FSX was ideally suited for this application, together with the iSTAR plates, which performed comparable to similar available microplates.