Introduction
Peatland ecosystems represent a significant global store of carbon (C) due to the historic accumulation of organic matter resulting from suppressed microbial decomposition. Decomposition is limited by extracellular enzymes that are produced by the plant and microbial communities and function independently of the microbial community. The storage of carbon in peatland ecosystems is therefore regulated in part by the environmental factors that determine extracellular enzyme activities. Understanding the environmental controls and processes that regulate extracellular enzyme activities is therefore critical in determining the fate of carbon in peatland ecosystems. Here we present the results from a method developed to measure potential extracellular enzyme activities in peat using a BMG LABTECH microplate reader. The assay is based on the use of fluorogenic methylumbelliferyl (MUF) substrates that fluoresce upon enzymatic cleavage allowing the amount of product to be measured.
Materials & Methods
- Black 96-well microplates, F-bottom, Greiner Bio-One
- MUF and MUF substrates, Sigma-Aldrich
- BMG LABTECH microplate reader
Peat samples were collected from Langdon moor, Durham, UK in order to determine the concentration of methylumbelliferyl (MUF) substrates to be added to peat solutions to achieve enzyme ‘active-site’ saturation and the optimal assay incubation length.
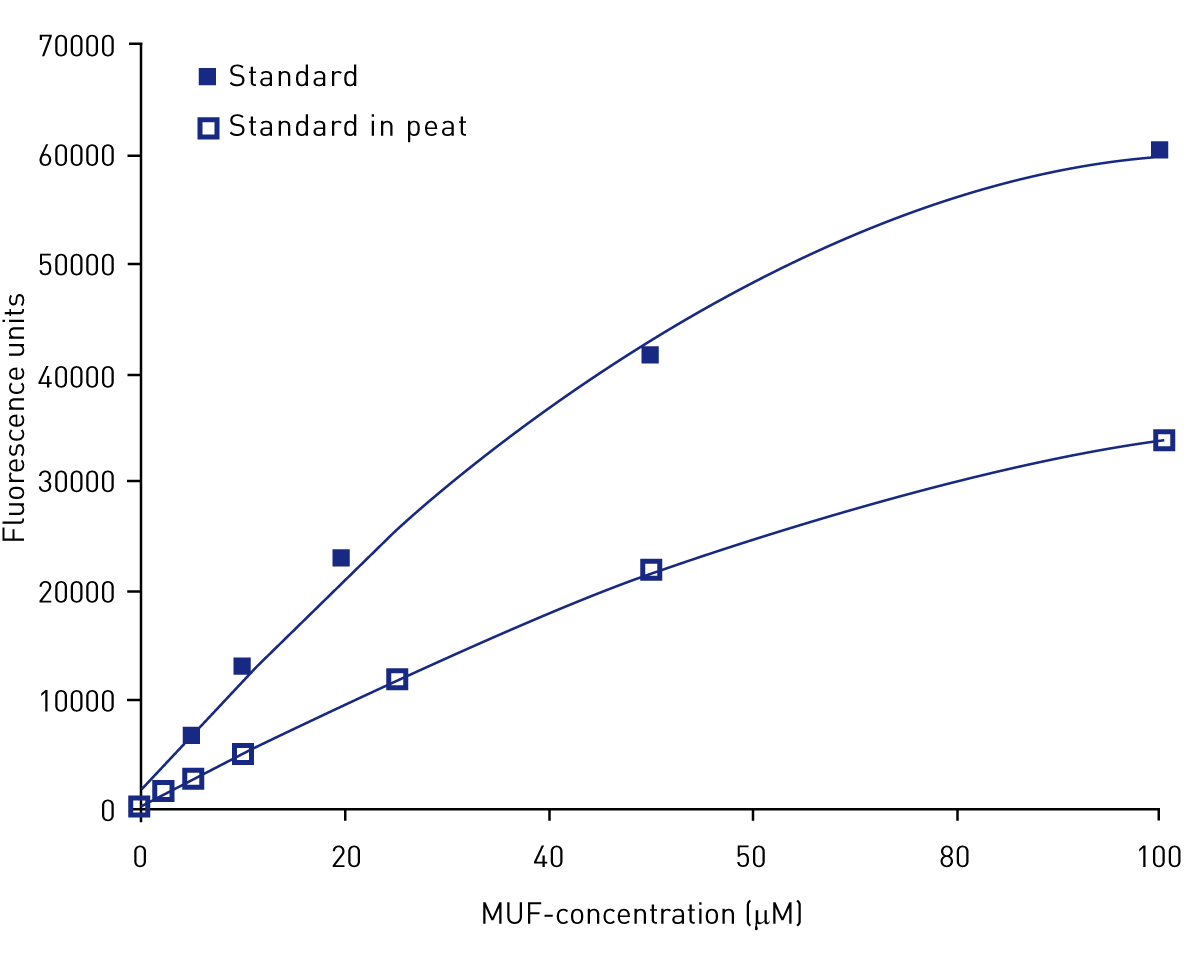
The hydrolytic enzymes β-glucosidase, cellobiohydrolase, and N-acetylglucosaminidase (or chitinase) were determined using MUF artificial substrates 3 (see Table 1). For each peat replicate sample, ten cm3 of peat was placed in a 50 mL centrifuge vial and deionised water added up to the 50 mL mark. The vial was shaken by hand for 30 seconds and mixed using a vortex for a further 30 seconds. Using a sawn-off pipette tip, 0.75 mL of peat slurry was placed into a 1.7 mL centrifuge vial. To the vial, 0.75 mL of MUF substrate was added and samples were incubated at field temperature for one hour. Samples were centrifuged for 5 minutes at 12,000 rpm and 300 μL of supernatant transferred to a well in a plate. Fluorescence was determined on a BMG LABTECH microplate reader at 330 nm excitation and 450 nm emission wavelength. Enzyme activities were determined from the fluorescence units using a standard calibration curve of methylumbelliferone (MUF) and expressed as rates of MUF production (μmol MUF per g-1 dry peat weight per min-1). Fluorescence quenching is a potentially interfering process which decreases the intensity of the fluorescence emission and occurs in water containing peat-derived compounds. The standard calibration curve accounted for quenching by dissolving the MUF standard in 150 μL of centrifuged peat slurry for each peat replicate.
Table 1: Enzymes and enzyme MUF substrates
| Enzyme | MUF substrate | Reaction |
| Cellobiohydrolase | MUF-β-Dcellobioside | Cellulose polymers to dimers |
| β-glucosidase |
MUF-β-Dglucopyranoside
|
Cellulose dimers to monomers
|
| Chitinase |
MUF-N-acetyl-β-Dglucosaminide
|
Chitin to acetylglucosamine
|
The assay above was performed with substrate concentrations varying from 0 to 500 μM in order to determine the optimal substrate concentration. Using the optimal substrate concentrations, samples were then incubated for varying lengths of time to determine the optimal assay incubation length.
Results & Discussion
Figure 1 shows the effect of increasing substrate concentration on the activity of cellobiohydrolase. Activity followed Michaelis-Menten kinetics and reached saturation at approximately 200 μM of substrate.
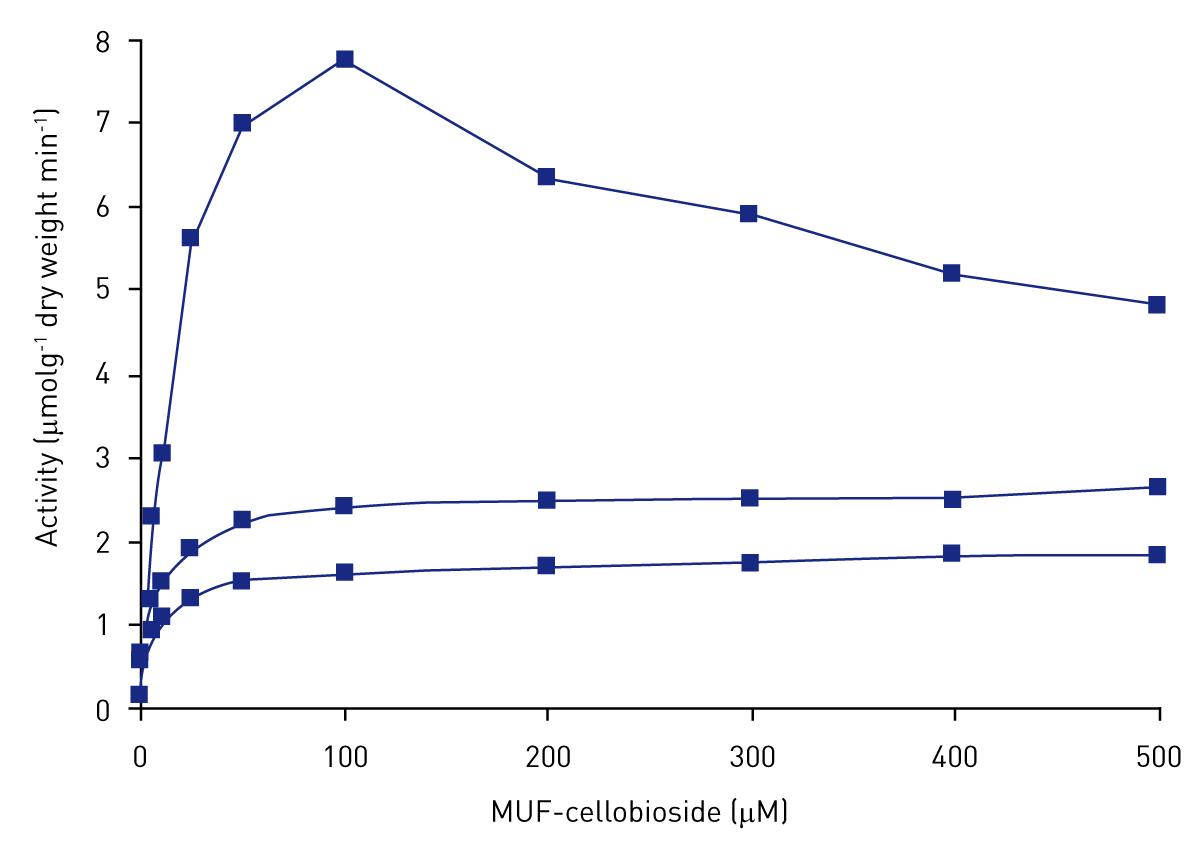
One of the replicates exhibited substrate inhibition at 100 μM of substrate resulting in a relative standard deviation of 85 % at 100 μM of substrate. This is mainly due to spatial variation of enzyme activity within the block of peat being analysed, NOT analytical variation.
Figure 2 shows the effect of increasing substrate concentration on the activity of β-glucosidase. Activity followed Michaelis-Menten kinetics and reached saturation at approximately 200 μM of substrate. One of the replicates exhibited substrate inhibition at 100 μM of substrate resulting in a relative standard deviation of 91 % at 100 μM of substrate.
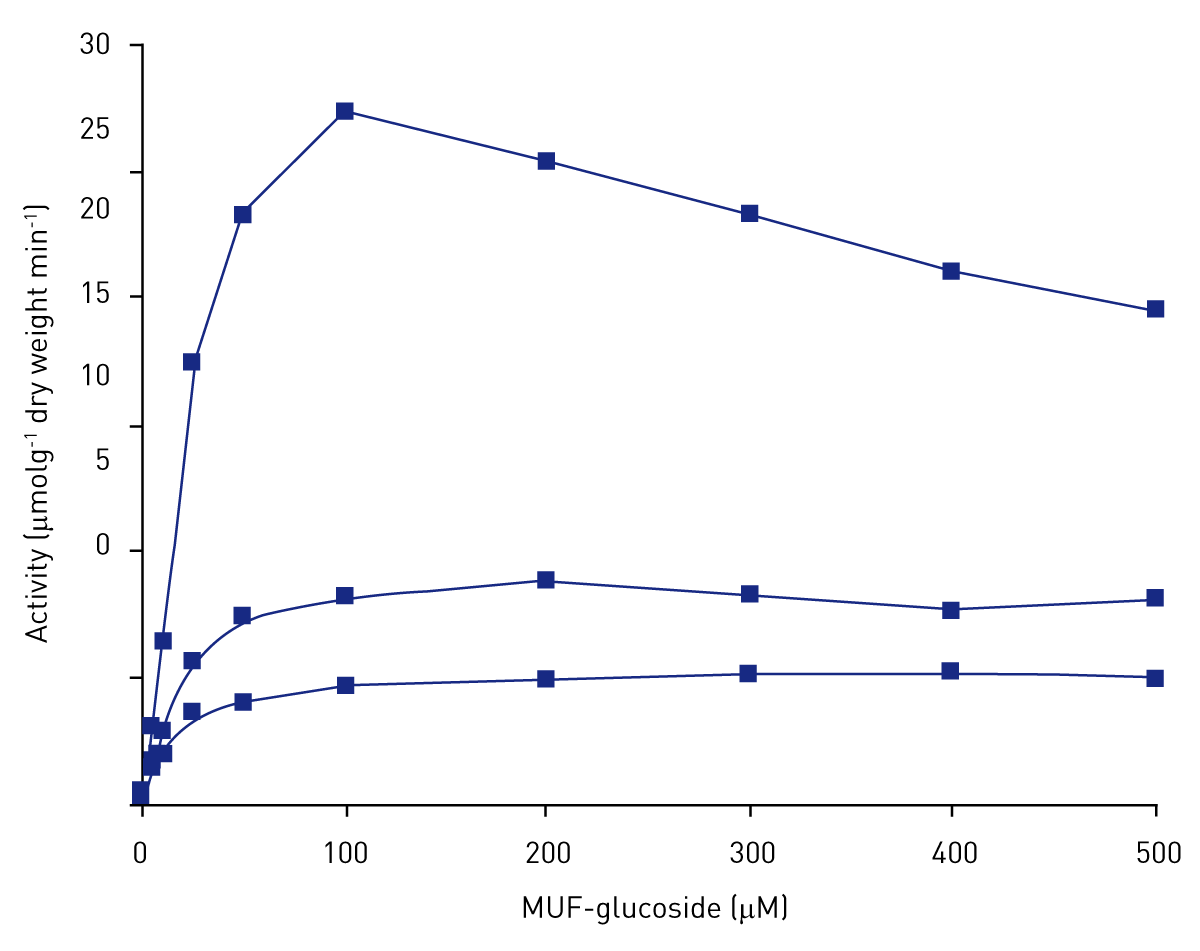
Figure 3 shows the effect of increasing substrate concentration on the activity of chitinase. Activity followed Michealis-Menten kinetics and reached saturation at approximately 300 μM of substrate.
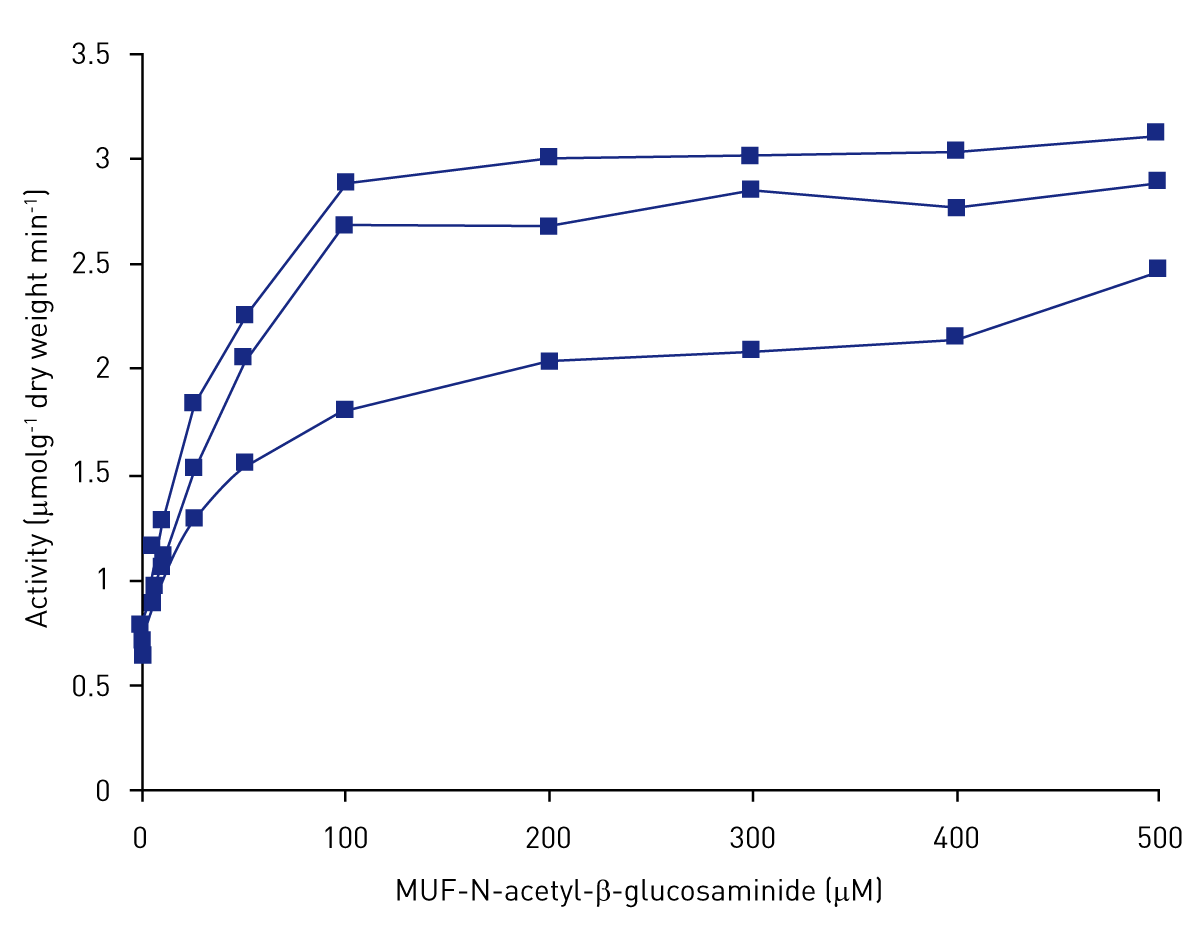
Figure 4 shows the significant quenching effect of peat on fluorescence.
 Time course incubations performed using optimal substrate concentrations for cellobiohydrolase, β-glucosidase and chitinase showed that one hour was the maximum amount of time that reaction rates remained linear (data not shown).
Time course incubations performed using optimal substrate concentrations for cellobiohydrolase, β-glucosidase and chitinase showed that one hour was the maximum amount of time that reaction rates remained linear (data not shown).
Conclusion
The extracellular enzymes β-glucosidase, cellobio-hydrolase and N-acetylglucosaminidase reached substrate saturation at 200 μM, 200 μM, and 300 μM of MUF respectively. Measured enzyme activities were linear for up to 120 minutes although 60 minutes duration of incubation was chosen for future applications to minimise variation between replicates. One replicate exhibited substrate inhibition for the β-glucosidase and cellobio-hydrolase assays.
The results show that potential extracellular enzyme activities can be determined in peat at low cost and within a short period of time using a BMG LABTECH microplate reader.