Introduction
Membrane fluidity and other properties of lipids are commonly assessed using various fluorogenic membrane probes and fluorescence polarization (FP) measurements. The basic principle is that alterations in lipid packing (e.g. temperature-dependent lipid phase transitions) change the mobility of a membrane-bound fluorophore. The latter parameter (specifically, “rotational relaxation”) can be monitored by exciting the fluorophore with a polarized light and measuring the emitted light in two planes - parallel and perpendicular to the polarization plane of the excitation light. FP is defined as the following ratio: III – Iꓕ / III + Iꓕ , where III and Iꓕ are fluorescent intensities measured in the parallel and perpendicular channels, respectively.
Membrane fluidization increases mobility of the dye and decreases the intensity of the emitted parallel component. Accordingly, FP is reciprocally related to the membrane fluidity. Optical characteristics of the fluorescent dye DPH strongly depend on the environment; the dye is almost non-fluorescent in aqueous solutions, while binding to the hydrophobic region of the membrane results in a sharp increase in the fluorescence signal (with an excitation maximum in UV range).
This note describes the use of a BMG LABTECH microplate reader for measurements of relative changes in lipid fluidity of DPH-labeled membranes.
Materials & Methods
- Phospholipids from Avanti Polar Lipids
- Mitochondria isolated from rat liver
- Opaque-walled, clear bottom 96-well plates from Costar
- Microplate reader from BMG LABTECH, equipped with UV FP optics
Membrane fluidity measurements were performed in liposomes and isolated mitochondria. 100 µl of samples were placed in a microplate. A 355 nm filter is used for excitation. Emission is measured at 430 nm.
To validate the method, FP measurements were taken using liposomes with defined gel-to-liquid crystalline phase transition temperature. Unilamellar liposomes were prepared from 1-myristoyl-2-palmitoylsn-glycero-3-phosphocholine (MPPC). The transition temperature for this lipid is in the 35 - 37°C range. For fluorescent labeling, MPPC liposomes were pre-incubated with 10 µM DPH (Sigma) at 45°C for 30 min in a KCl-based buffer (150 mM KCl; 10 mM HEPES, pH 7.4; 2 mM EGTA). Steady-state FP measurements were taken at 25 - 45°C. Temperature on the microplate reader’s incubator was increased by 2°C and FP measurements were taken after equilibrating the samples for 10-15 minutes. Data were acquired in endpoint mode.
A more complex model was investigated, native mitochondrial membranes. Isolated rat liver mitochondria (0.2 mg/ml) were preincubated with 10 µM DPH at room temperature for 40 minutes. Temperature in the microplate reader’s incubator was increased by 2°C and the samples were equilibrated for 10 minutes at each given temperature before FP measurements were taken. Mitochondria used in these experiments were in a deenergized (non-respiring) state. Lastly, the effects of membrane active drugs dibucaine and propranolol on membrane fluidity in liposomes and mitochondria were determined.
Liposomes were formed from the following lipid mixture mimicking mitochondrial membranes: 47% phosphatidylcholine (PC), 28% phosphatidylethanolamine (PE), 9% phosphatidylinositol, 9% phosphatidylserine (PS), and 7% cardiolipin (CL). Data were acquired in kinetic mode.
Results & Discussion
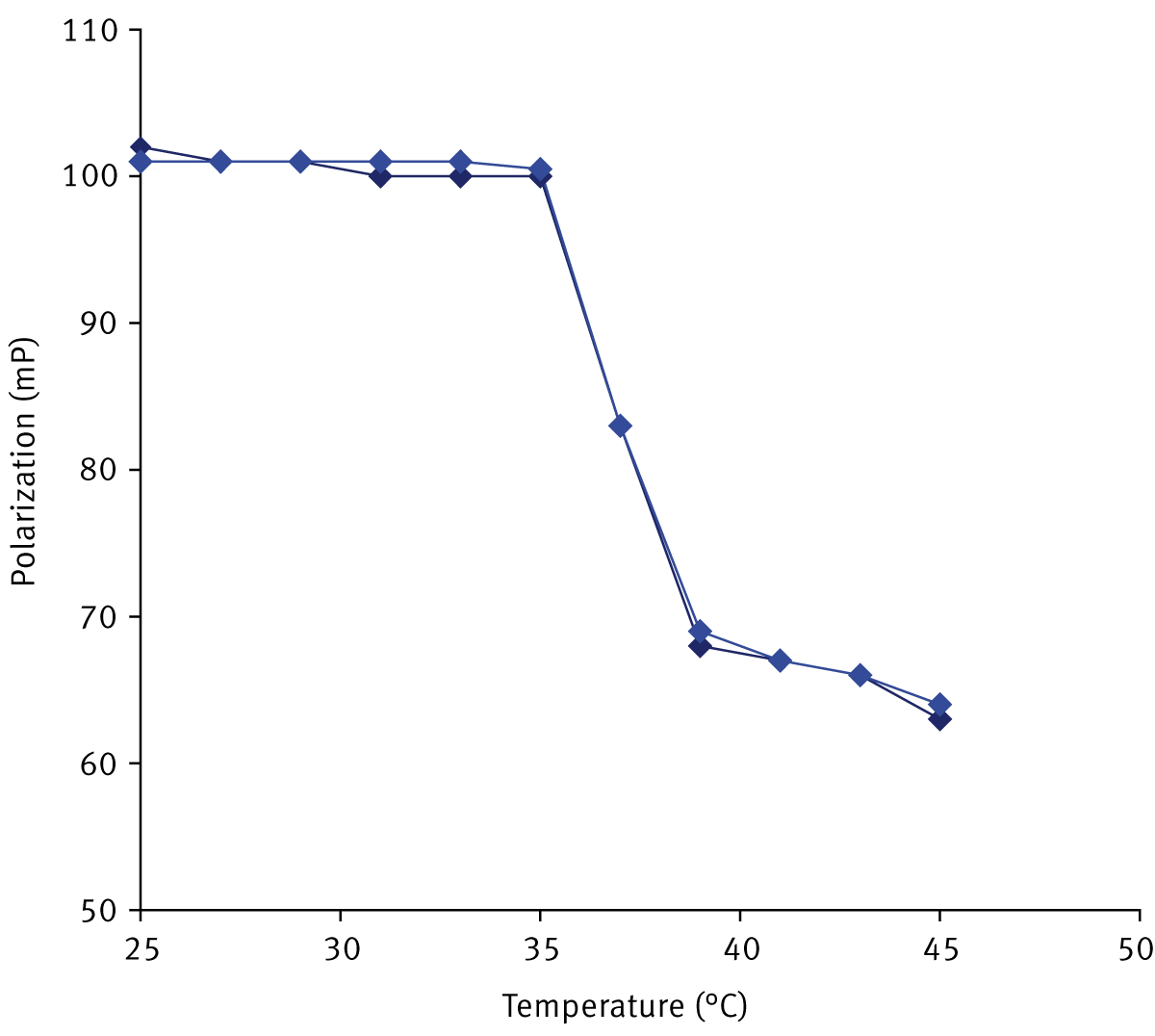
As shown in Figure 1, increasing the temperature from 25 to 35°C did not change the FP values in MPPC liposomes. This reflects the expected high ordered gel state of the lipid in this temperature range. Increasing the temperature from 35 to 39°C results in a sharp, large-scale decrease in FP values corresponding to the phase transition. In the liquid phase, FP values continued to decline as the temperature increased to 45°C.
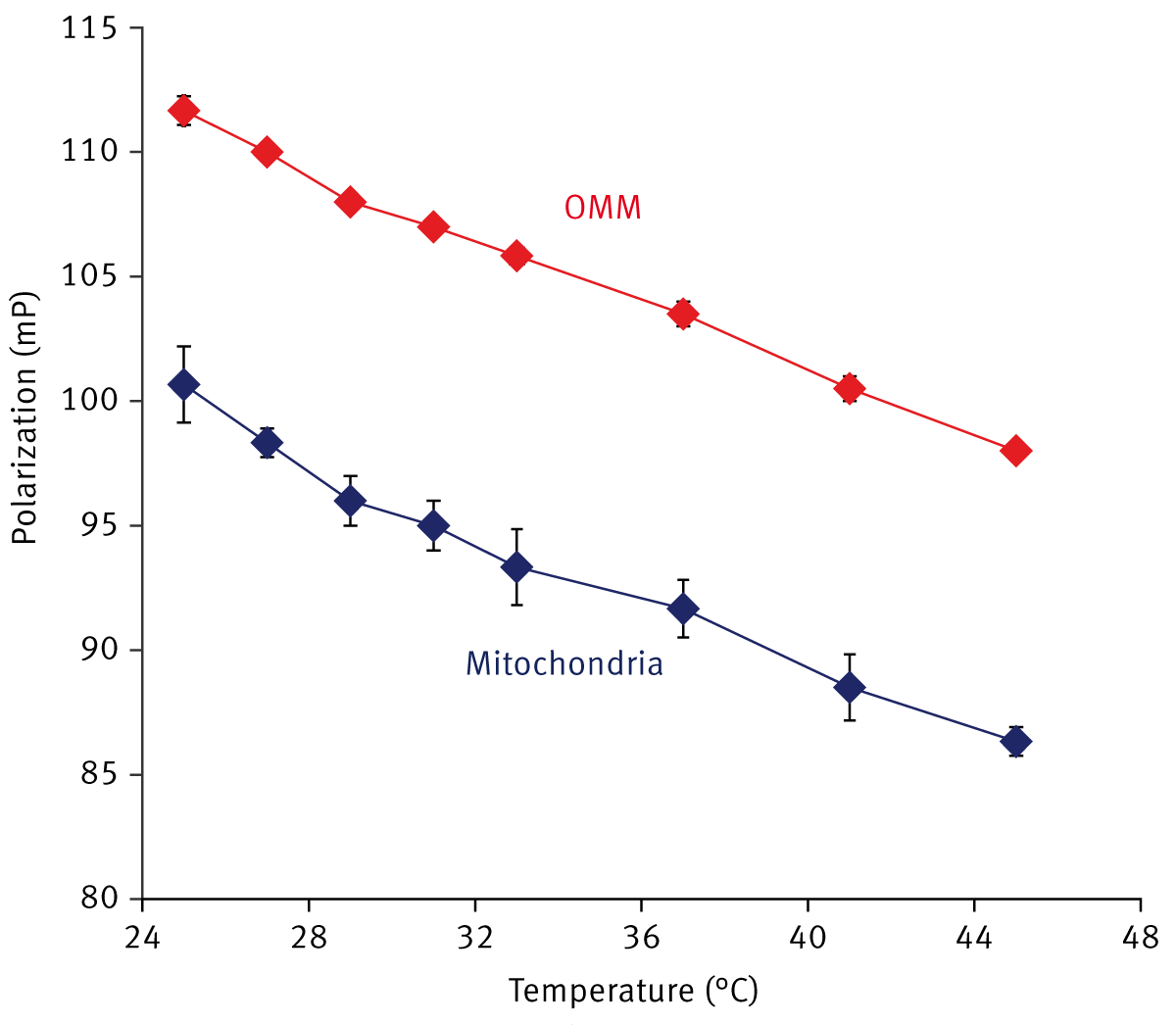
Unlike MPPC vesicles, no rigid (temperature-insensitive) state or large-scale phase transitions were observed in the mitochondrial membranes (Fig. 2). A gradual decrease in FP in the 25 - 45°C temperature range is seen, which is consistent with previously published data on membrane fluidity measurements in DPH-labeled deenergized mitochondria. Relative FP values in whole mitochondria and purified outer mitochondrial membranes (OMM) were also compared. The patterns of temperature-dependence were similar in both membrane systems, but the relative FP values in OMM were significantly higher than those obtained in whole mitochondria (Fig. 2). This result can be readily explained by the higher cholesterol level in the OMM compared to the inner mitochondrial membrane.
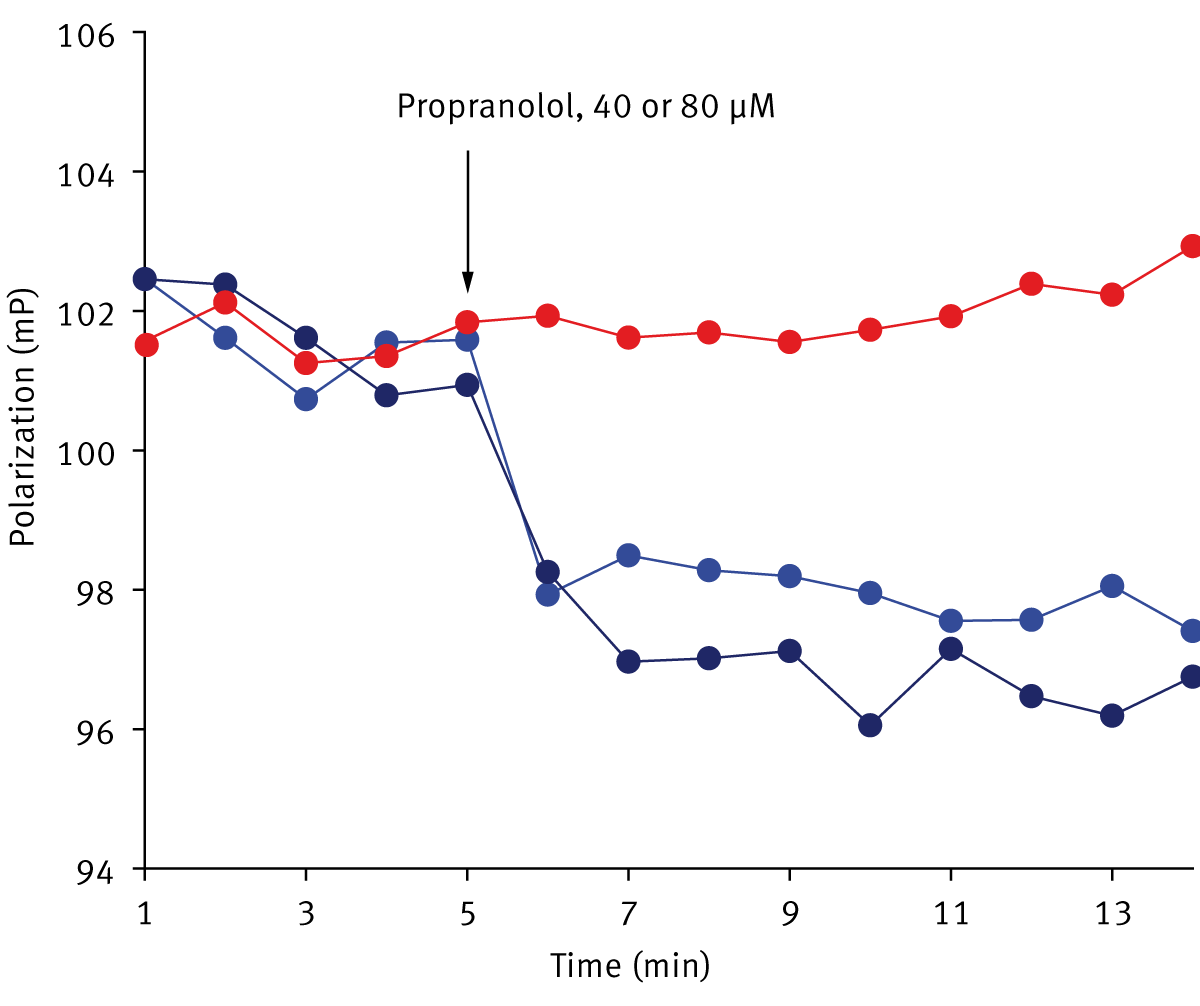
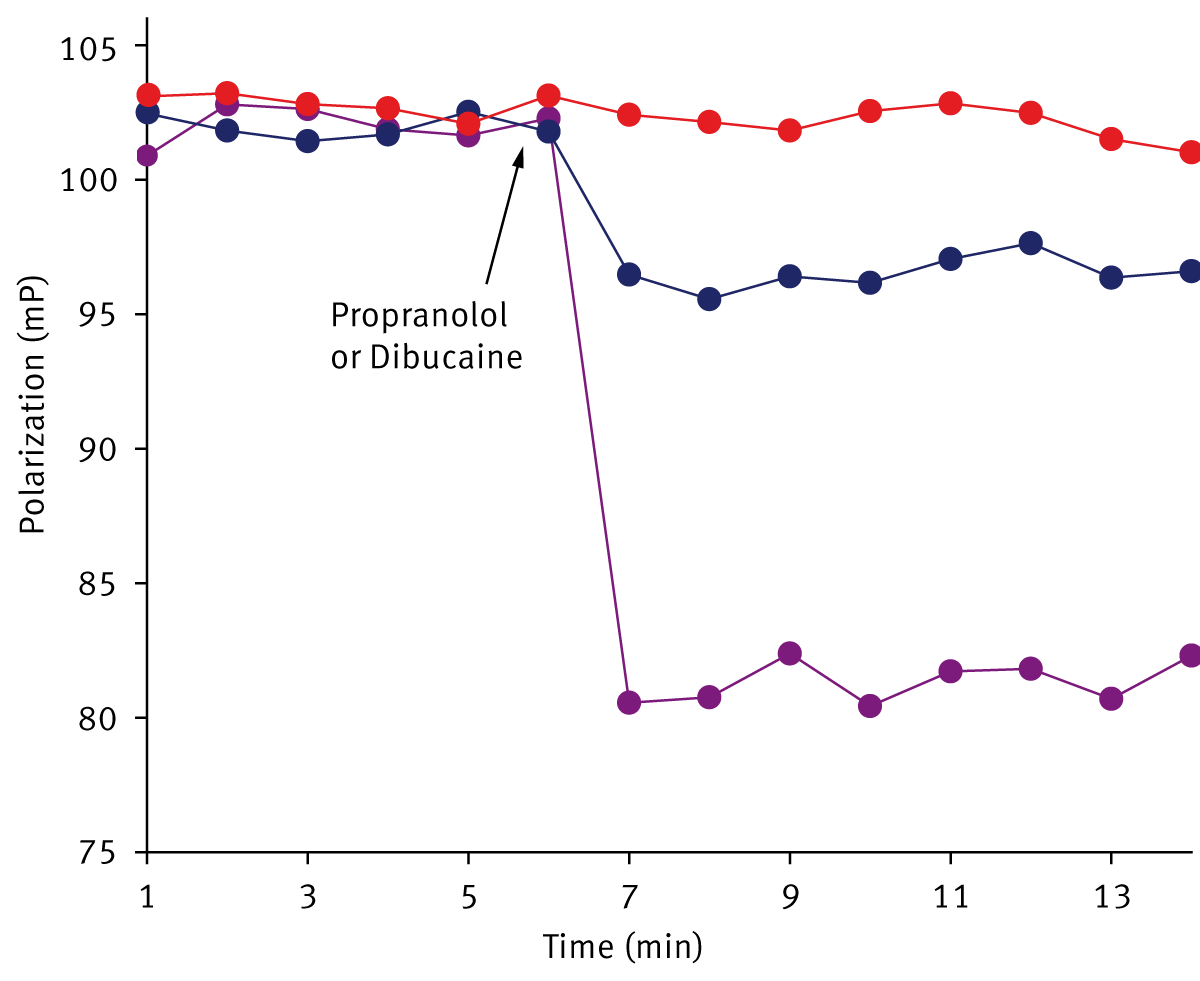
Finally, the effects of propranolol and dibucaine on liposomes and isolated mitochondria were examined. Depending on concentrations, these drugs can increase membrane fluidity or rigidify some membranes. Figure 3 shows that the addition of propranolol to liposomes (panel A) results in a rapid decrease in relative FP values. A similar effect was observed in isolated mitochondria (panel B). The propranolol-induced decrease in FP was smaller in comparison to the dibucaine induced changes, but nonetheless was readily detected. The concentrations of the drugs (40-80 µM) were in the same range as those affecting mitochondrial activities. The data suggest that some of the effects of the drugs on mitochondrial function may be due to an increase in membrane fluidity.
Conclusion
This study demonstrates the suitability of the BMG LABTECH microplate reader with UV FP optics for measurements on membrane fluidity in a microplate-based format. The method allows for the monitoring of large scale alterations in membrane fluidity (such as phase transitions), as well as more subtle changes in lipid dynamics. This approach can be used as a high-throughput screening assay in the search for compounds altering lipid fluidity.