Introduction
Small G-proteins or small GTPases are a family of nearly 200 proteins in humans that can bind and hydrolyze guanosine triphosphate (GTP). They are considered active when they are bound to GTP and inactive when bound to GDP. They signal to a wide variety of effector proteins and are involved in the regulation of cellular processes such as growth and cell movement. Dysregulated small G-protein function has been associated with many diseases, including cancer1.
The best characterized small G-proteins belong to the Ras superfamily which is further sub-divided into 5 groups which include the Rho GTPase family. Rho GTPase family members are involved in dynamic regulation of the cell cytoskeleton and contribute to cell movement and cancer progression2.
Foerster´s Resonance Energy Transfer (FRET) biosensors have proved to be a valuable tool in the study of Rho GTPases3. But since the family members are known to act together being able to observe their actions simultaneously is vital to our understanding4. Here we describe a FRET biosensor for Rac1 that uses near-infrared (NIR) fluorescent proteins and thus is spectrally resolved from existing biosensors that are based in the blue/green/yellow spectral range while also being less susceptible to autofluorescence issues often observed with cell-based samples3. The spectral scanning capabilities of the CLARIOstar Plus proved useful in monitoring the function of these biosensors in a multiplexing approach.
Assay principle
Figure 1 shows a diagram of the NIR FRET Rac1 biosensor3. It takes advantage of the molecular switch nature of small G-proteins. In the active GTP-bound state binding of the p-21 binding domain (PBD), derived from p21 activated kinase 1, to Rac1-GTP occurs with high affinity. The biosensor incorporates a second PBD that has GTPase-binding deficient mutations (figure 1; depicted in red). This serves as an auto-inhibitory motif to limit FRET competent interactions to the presence of active GTP-bound Rac1.
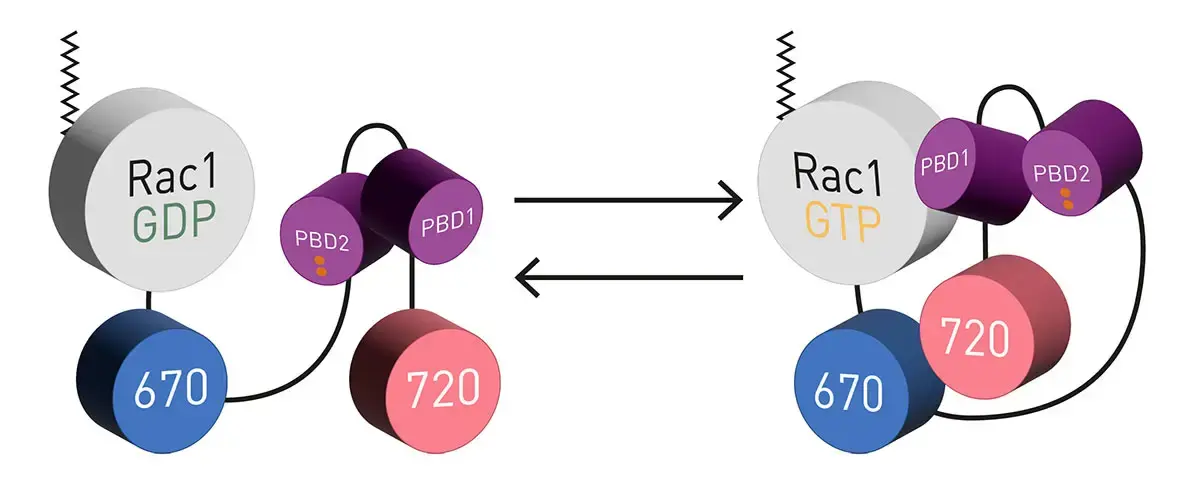
The biosensors use miRFP670 and miRFP720 as FRET pair attached to the C-terminus of Rac1. Two PBD’s provide appropriately tuned interaction with Rac1 (Adapted from Shcherbachova et al.)3.
Materials & methods
- 12-well, clear, TC-treated, microplates (Costar)
- CLARIOstar Plus with red extended PMT
- Other chemicals and reagents were obtained from commercial sources
Experimental Procedure
HEK293 cells were transfected with plasmids carrying the indicated Rac1 WT or Rac1 mutant biosensors. 400 ng of biosensor plasmid was used in 100 µL OptiMEM solution (all DNA amount listed here are optimized for 6-well plate format. For 12-well plate format, 50% of this mixture was then used to setup technical duplicates).
Where indicated, cells were additionally transfected with modifying proteins, 400 ng of these plasmids were used except for 4x GDI (1600 ng). Empty plasmid control DNA was added to a total of 2 µg of DNA as needed. Transfections were performed using the PEI reagent5, and the transfection mixture volume and the cell culture media volume per well and the cell seeding densities were appropriately optimized for the 12-well plate format. Adherent cells were read in clear L15 media with 10% FBS at room temperature.
Instrument settings
|
Fluorescence Intensity, spectrum
|
||
|
Optic settings
|
Excitation wavelength (nm) |
591-52
|
| Emission wavelengths (nm) |
640-15 to 802-15
|
|
| Gain |
2600 |
|
|
Focal height
|
4.8 |
|
|
General settings
|
Flashes
|
60 |
|
Settling time (s)
|
0.5 |
|
Results & Discussion
Spectral data were blank corrected using spectra from HEK293 cells transfected with control empty plasmid. The data were then scaled using the RFU at 676 nm as 100%. Representative data are shown in figure 2.
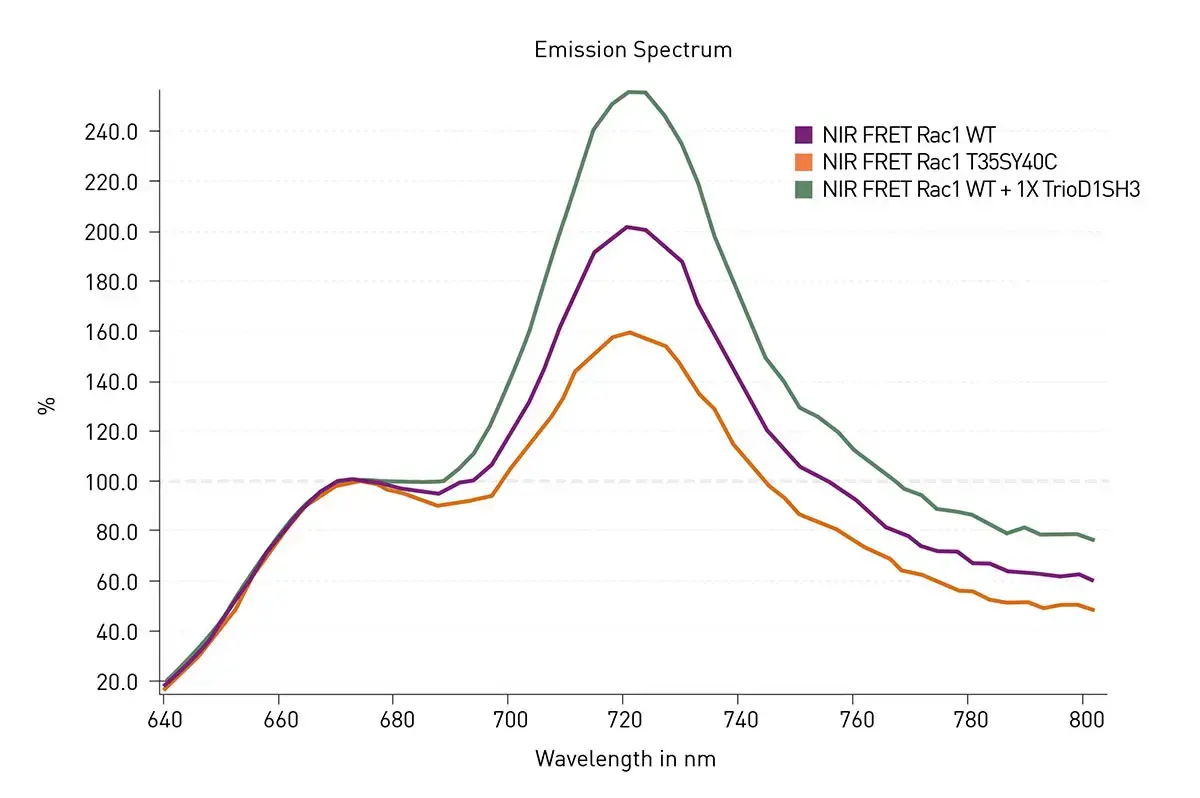
As expected for the conditions used WT Rac1 exhibited FRET signal transfer due to Rac1 activation resulting in a signal increase at 721 nm. The extent of FRET was decreased in T35S-Y40C Rac1. Since this mutant should not bind the PBD built into the sensor, the response agrees with the expectations. Finally, the expected increase in FRET could be seen upon the addition of an activator protein fragment (TrioD1SH3) which was co-transfected with WT NIR-Rac1. The experiments shown in figure 2 were repeated and additional mutations and co-transfections were tested (figure 3). The results in figure 3 related well to the expectations for mutations and co-transfections. Of note the Q61L exhibited a significant increase in FRET which agrees with this being the strongest activating mutation of Rac1. Three dominant negative mutants (T17N, G15A and D118A) showed a moderate increase in FRET signal, indicating that the stable interaction with the upstream activators elevates FRET. Effector binding mutations (T35SY40C and T35S) showed reduced FRET. In addition, co-transfection of GDI at 4x excess, a negative inhibitor protein that binds and deactivates GTPase, significantly reduced WT FRET. Addition of the activator fragment of Rac1 (TrioD1SH3) to the WT biosensor resulted in an increase in FRET signal while the same fragment but with catalytically deactivating mutations (TrioD1SH3-DN) had no effect on the WT FRET.
The results in figure 3 related well to the expectations for mutations and co-transfections. Of note the Q61L exhibited a significant increase in FRET which agrees with this being the strongest activating mutation of Rac1. Three dominant negative mutants (T17N, G15A and D118A) showed a moderate increase in FRET signal, indicating that the stable interaction with the upstream activators elevates FRET. Effector binding mutations (T35SY40C and T35S) showed reduced FRET. In addition, co-transfection of GDI at 4x excess, a negative inhibitor protein that binds and deactivates GTPase, significantly reduced WT FRET. Addition of the activator fragment of Rac1 (TrioD1SH3) to the WT biosensor resulted in an increase in FRET signal while the same fragment but with catalytically deactivating mutations (TrioD1SH3-DN) had no effect on the WT FRET.
Conclusion
We show the validation of the NIR FRET Rac1 biosensors as a viable tool to detect changes in Rac1 activation. Mutations of Rac1 or co-expression of modifying proteins had the expected effect on FRET as assessed by spectral scanning using the CLARIOstar Plus. This was achieved using the Linear Variable Filter (LVF) monochromator that offers flexibility and filter-like sensitivity. These biosensors will be useful tools in furthering our understanding of complex small G-protein signalling..
References
- Yin, G., et al. Targeting small GTPases: emerging grasps on previously untamable targets, pioneered by KRAS. Sig Transduct Target Ther (2023) 8: 212
- Vega, F.M., Ridley, A.J. Rho GTPases in cancer cell biology. FEBS Lett (2008) 582: 2093-2101
- Shcherbakova, D.M., et al. Direct multiplex imaging and optogenetics of Rho GTPase enabled by near-infrared FRET. Nat. Chem Biol (2018) 14: 591-600
- Machacek, M. et al. Coordination of Rho GTPase activities during cell protusion. Nature (2009) 461: 99-103
- Ehrhardt, E., et al. Polyethylenimine, a cost-effective transfection reagent. Signal Transduct (2006) 6: 179-184